4 portions of Overnight Oats with Apple and Dried Fruit Prep:10 min + overnight resting
Ingredients
180 g (6 oz) rolled oats (Whole)
500 ml (2 cups) milk, Whole fat or your favorite homemade vegetable milk
30 ml (2 Tbsp) brown sugar (raw sugar or honey)
2 apples, finely diced
30 ml (2 Tbsp) finely sliced almonds or nuts
30 ml (2 Tbsp) sultana raisins or sliced prunes
30 ml (2 Tbsp) dried apricots, figs or dates, chopped
Instructions
Soak the oats in the milk.
Stir in all remaining ingredients. Mix well, cover with plastic wrap, and refrigerate overnight.
Serve with extra milk, cream or yogurt.
Benefits
Overnight oats are becoming a source of resistant starch, a type of soluble fiber that feeds a healthy gut microbiome, specifically the Akkermansia muciniphila species.
Akkermansia plays an important role in maintaining good gut health, including cancer prevention.
All the other ingredients provide protein, high-quality fats, fiber, vitamins, minerals, polyphenols, antioxidants, etc.
Keep the sugar at the “sweet spot” – in the middle to low.
While in Romania, I bought this amazing book: “Microbiomul: Sănătatea Începe în Intestin” by Prof. Dr. Maria Rescigno, in a Romanian translation of the original Italian book “Microbiota Geniale: Curare l’intestino per guarire la mente” (2023). I find the content very well explained to laypeople. The book is a fountain of new information that should interest all of us. The gut microbiome is considered a new organ that stays at the basis of human health. It is known that about 90% of all diseases start from gut microbiome dysbiosis. This book is available in English
The author, Dr. Maria Rescigno, PhD, a renowned biologist and researcher at Humanitas University, is recognized as one of the world’s leading experts on the microbiome. She has published over 200 papers in top journals like *Science* and *Nature*, and her groundbreaking discoveries include the gut vascular barrier (GVB) in 2015 and the plexal vascular barrier (PVB) in the brain in 2021. The book explores the intricate connections between the gut microbiome, the immune system, and the brain, emphasizing how modulating the microbiome can prevent and treat various health issues, particularly neurological and mood disorders.
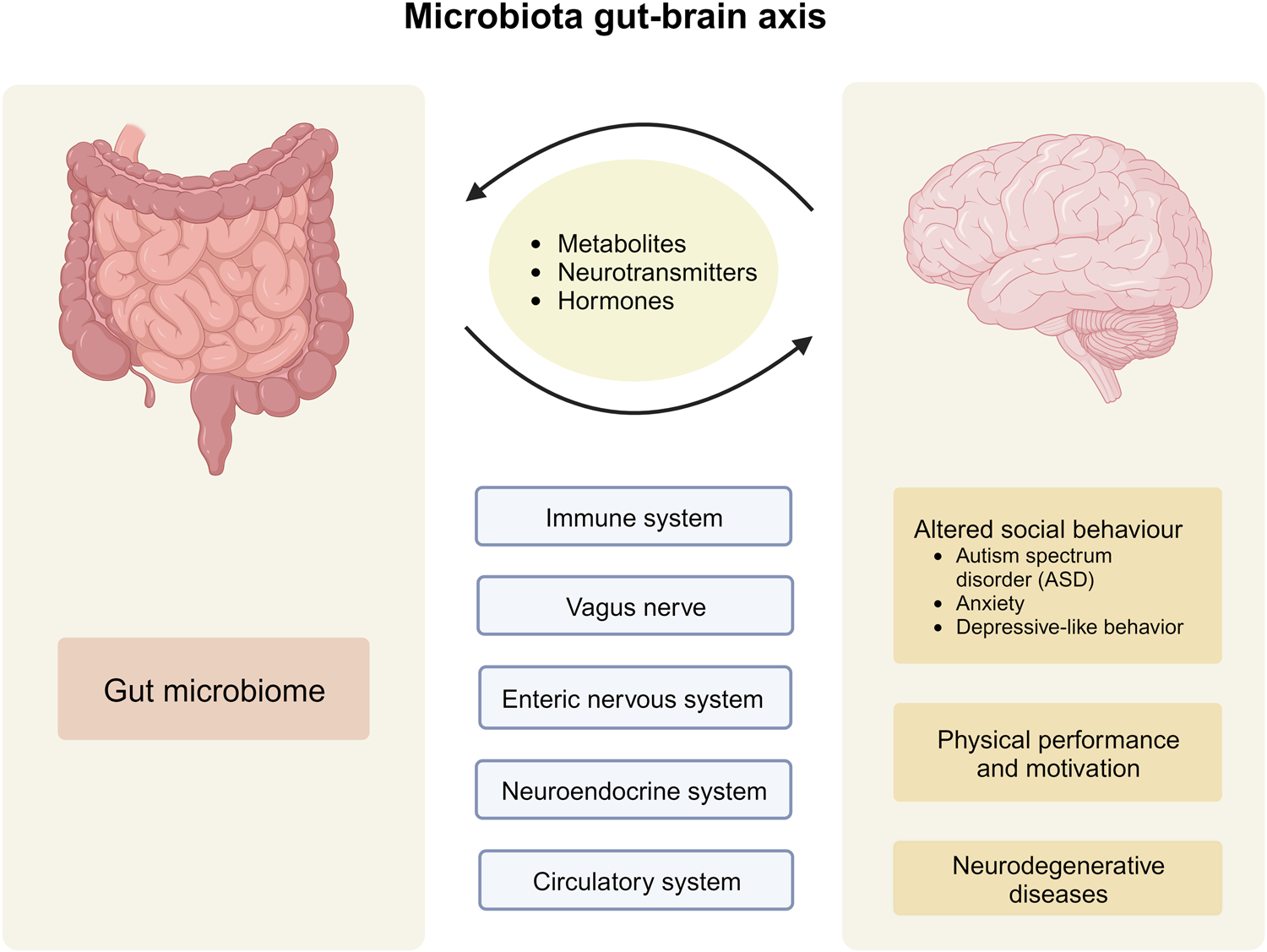
The central thesis is that health begins in the gut, where a rich population of microorganisms (the microbiome) aids digestion, communicates with the immune and nervous systems, and influences overall well-being. Rescigno highlights the bidirectional gut-brain axis, regulated by the microbiome, and draws from recent research—including her own—to show how imbalances (dysbiosis) in the gut can contribute to conditions like anxiety, depression, Alzheimer’s, Parkinson’s, autism, and eating disorders. The book combines scientific rigor with accessible explanations, offering practical strategies for microbiome modulation.
Chapter 1: A Brief Excursion into the World of the Microbiome
This chapter introduces the microbiome as an “additional organ” composed of bacteria, viruses, fungi, and other microbes inhabiting our mucous membranes, particularly the gut. Rescigno explains its evolution from being called “bacterial flora” to “microbiome,” detailing its roles in metabolism, immune modulation, and communication with the body. She describes the microbiome’s diversity, likening the gut to “guest rooms” or “shores” where microbes interact with host cells. The chapter sets the stage for understanding how this microbial world impacts health beyond digestion.
Chapter 2: What Are Barriers and What Role Do They Play?
Rescigno delves into protective barriers in the body, focusing on the intestinal vascular barrier (GVB), which she discovered, and its similarity to the blood-brain barrier. She discusses intestinal permeability and leaky gut syndrome (LGS), where breaches allow harmful substances to enter the bloodstream. The chapter also covers the two brain barriers: the blood-brain barrier and the newly identified plexal vascular barrier (PVB) in the choroid plexus, which controls substances entering cerebrospinal fluid. Drawing on experiments, including those by researcher Michal Schwartz, Rescigno explains how these barriers open and close in response to gut-derived stimuli, forming a vascular gut-brain axis.
Chapter 3: The Two Brains
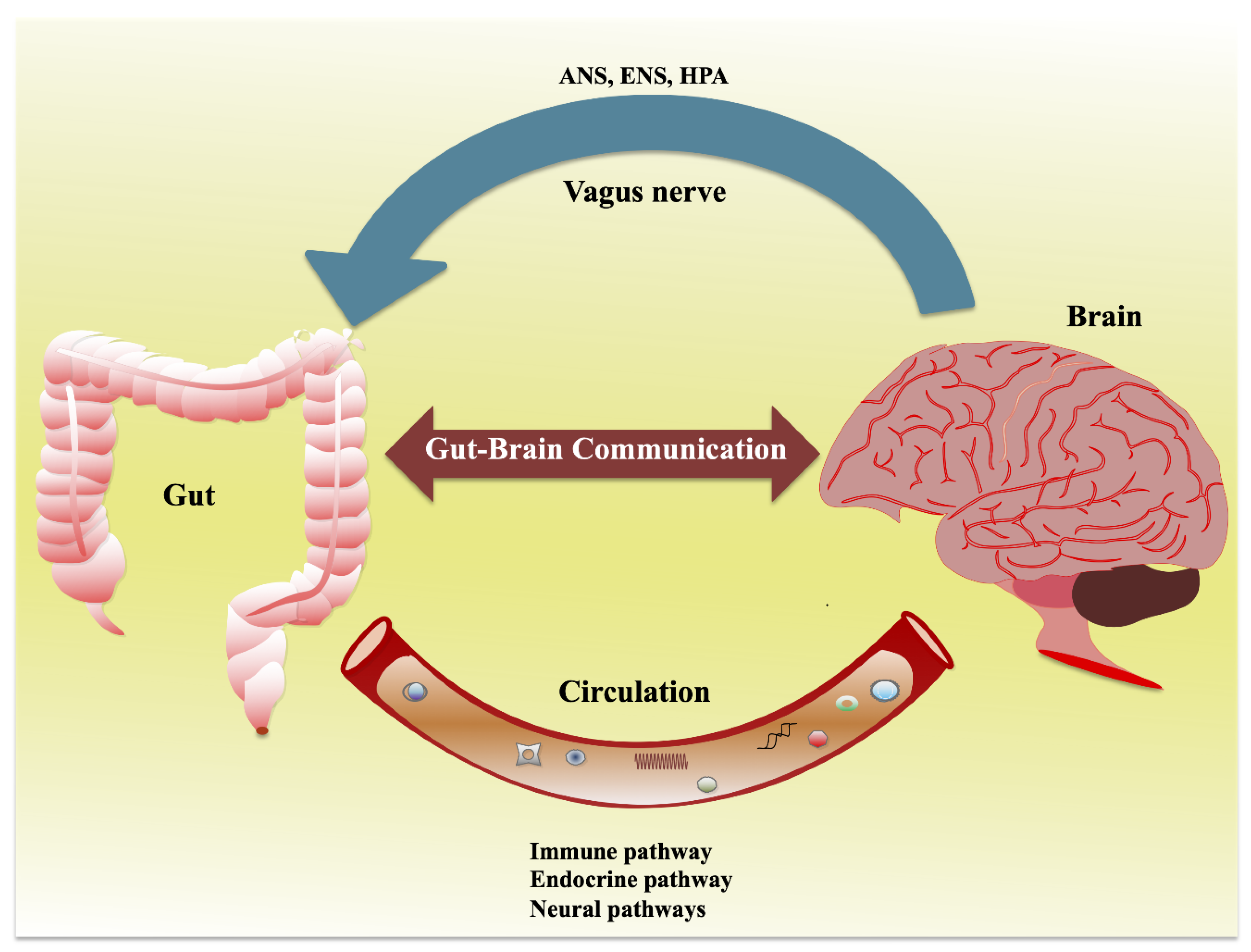
Here, the author explores the “second brain”—the enteric nervous system in the gut—which engages in constant dialogue with the central nervous system. Communication channels include the nervous system, vagus nerve, immune pathways, and hormonal signals. Rescigno details how microbes produce neurotransmitters and metabolites that influence mood control centers in the brain, acting as key players in this bidirectional exchange. She emphasizes that gut microbes can “write” messages that affect emotions and behavior.
Chapter 4: Neurological Diseases and the Microbiome
This extensive chapter links microbiome dysbiosis to various conditions. It questions causality (“chicken or egg?”) and examines microbiome changes in aging, Alzheimer’s (involving microglial cells), Parkinson’s (gram-positive/negative bacteria), sleep issues (melatonin), ALS, multiple sclerosis, IBS, anxiety (including sepsis-related), depression (major and bipolar), schizophrenia, psychosis, and childhood/adolescent disorders like autism (role of bifidobacteria) and drug-related psychosis risks. Beyond the gut-brain axis, it touches on periodontitis and neurodegenerative links.
Chapter 5: The Microbiome and…
Focusing on eating behaviors, this chapter discusses satiety, eating disorders like anorexia, and how diet influences social behavior via the microbiome. Rescigno presents evidence from studies showing microbiome composition differs in patients with nutritional disorders, and maternal nutrition can shape offspring behavior through microbial mechanisms.
Chapter 6: Prevention and Treatment of Neurological Diseases Through Approaches That Modulate the Microbiome
Rescigno shifts to actionable strategies, covering nutrition’s protective role against neurodegeneration. She highlights plant-based antioxidants like polyphenols, flavonols for Parkinson’s, Korean red ginseng for Alzheimer’s, ketogenic diets for IBS, olive/camellia oils, and omega-3s. The chapter reviews pre-, pro-, and postbiotics, clinical trials (e.g., Lacticaseibacillus rhamnosus for mood in overweight patients, probiotic mixes for depression and autism), fecal microbiota transplants, bacterial metabolites, and fermented foods like kimchi. Preclinical models and in vitro studies support these interventions.
Chapter 7: Future Perspectives
The book addresses limitations in microbiome analysis and offers practical advice: avoid leaky gut and high animal fat diets; consume fiber-rich foods, healthy fats, Mediterranean diet staples (legumes + cereals); prefer fermented foods; consider in vitro fermentation tests and postbiotics; try a 15-day grain-free test; and incorporate daily exercise. Rescigno discusses open research challenges and potential advancements in biomedical applications.
Conclusions
Dr Maria Rescigno concludes that modulating the microbiome through lifestyle and targeted interventions can preserve intestinal barriers, balance the gut-brain axis, and mitigate neurological risks, empowering readers to take control of their health.
**Citation:** Rescigno, Maria. *Microbiomul: Sănătatea Începe în Intestin*. Translated by Donna Oprea, Philobia, 2024. (Original: *Microbiota Geniale: Curare l’intestino per guarire la mente*. Vallardi, 2023.)
Synbiotics are combinations of probiotics (live beneficial bacteria) and prebiotics (non-digestible fibers that feed them), designed to synergistically improve gut health.
They enhance SCFA production—primarily acetate, propionate, and butyrate—more effectively than probiotics or prebiotics alone by providing both the microbes and their preferred substrates for fermentation in the colon. This synergistic combination boosts microbial diversity, SCFA yields, and promotes overall health.
Mechanisms of Enhanced SCFA Production
Fermentation Synergy: Prebiotics like inulin or fructo-oligosaccharides (FOS) selectively nourish probiotic strains (e.g., Bifidobacterium, Lactobacillus), leading to increased breakdown of fibers into SCFAs. For instance, synbiotics can elevate butyrate (from butyrate-producing bacteria) and acetate levels without promoting harmful byproducts.
Microbiota Modulation: They shift the gut microbiome toward SCFA-producing species, reducing pH and inhibiting pathogens while optimizing mineral absorption and barrier function.
Dose and Formulation: Encapsulated synbiotics (e.g., Limosilactobacillus fermentum with prebiotics) survive digestion better, amplifying colonic fermentation.
Evidence from Recent Studies
A 2025 meta-analysis of 28 RCTs (randomized control studies) found synbiotics significantly increased fecal SCFAs (e.g., acetate +15%, butyrate +20%) and improved microbiota composition in adults with metabolic disorders, outperforming single interventions.
In a 2023 preclinical trial, the synbiotic AG1® (probiotic blend + prebiotic fibers) raised total SCFAs by 25-30%, including propionate, in simulated gut models, linking to anti-inflammatory effects.
A 2024 double-blind RCT showed synbiotic intake (probiotics + FOS) enhanced carbohydrate metabolism, boosting SCFA production by 18% and aiding blood sugar control.
Studies from 2023-2024 confirm that synbiotics reduce systemic inflammation via SCFAs, with meta-analyses reporting lowered CRP and IL-6 levels.
Health Implications Higher SCFA production from synbiotics supports gut integrity, immune modulation, metabolic health (e.g., insulin sensitivity), and reduced chronic disease risk like IBD or obesity.
For optimal results, incorporate via foods (e.g., yogurt with oats) or supplements, starting low to minimize bloating.
Consult a professional for tailored use.
Prebiotics vs. Probiotics: Key Differences and Benefits Both prebiotics and probiotics support gut health in complementary ways:
Probiotics introduce live beneficial bacteria, while prebiotics nourish existing ones.
Often combined as synbiotics for enhanced effects, they promote microbiome balance, which is linked to digestion, immunity, and more.
Below is a comparison based on recent expert guidance.
Aspect
Prebiotics
Probiotics
Definition
Non-digestible fibers (e.g., inulin, oligosaccharides) that feed beneficial gut bacteria, acting like “fertilizer” for the microbiome.
Live microorganisms (e.g., Lactobacillus, Bifidobacterium) that provide health benefits when consumed in sufficient amounts.
How They Work
Resist digestion in the upper gut, reaching the colon to selectively stimulate growth of good bacteria, helping them outcompete harmful ones.
Colonize the gut temporarily, producing beneficial compounds like SCFAs and modulating immune responses.
Food Sources
High-fiber plants: onions, garlic, leeks, asparagus, bananas (especially green), apples, oats, barley, chickpeas, flaxseeds. Also in supplements.
Fermented foods: yogurt, kefir, sauerkraut, kimchi, miso, kombucha, tempeh. Also in supplements and fortified foods.
Health Benefits
Improve digestion and regularity; reduce inflammation; support immune function; may aid weight management and blood sugar control by boosting SCFA production.
Enhance digestion (e.g., reduce IBS symptoms); strengthen immunity; decrease antibiotic-associated diarrhea; support mental health via gut-brain axis.
When to Choose
Ideal for individuals with a fiber-deficient diet;
Best for long-term microbiome support. Start low to avoid bloating.
Useful after antibiotics or for acute gut issues; choose strains targeted to needs (e.g., Lactobacillus for diarrhea).
Potential Drawbacks
May cause gas/bloating initially in high doses; not suitable for everyone (e.g., FODMAP-sensitive).
Variable efficacy by strain; some may cause mild side effects like gas; shelf life matters for live cultures.
For optimal results, incorporate both through a diverse, plant-rich diet.
Aim for 25–30g fiber daily for prebiotics alongside probiotic foods.
Consult a healthcare provider for supplements, especially with conditions like IBS.
Dietary Sources of Inulin and Fructo-Oligosaccharides (FOS)
Inulin and fructo-oligosaccharides (FOS) are types of prebiotic fibers naturally occurring in many plant-based foods, particularly those that store energy as fructans.
These compounds are found in varying concentrations (typically measured in grams per 100g of food) and can also be added to processed foods like cereals, breads, and snacks as ingredients labeled “inulin” or “FOS.” You should get them from the real foods. Avoid processed foods!Below is a table summarizing key natural dietary sources, based on reliable nutritional data.
Amounts are approximate and can vary by preparation (e.g., raw vs. cooked).
Food Source
Type (Inulin/FOS/Both)
Approximate Amount per 100g
Notes
Chicory Root
Inulin
35.7–47.6 g
Highest natural source; often used in supplements or coffee substitutes.
Jerusalem Artichoke
Inulin
16–20 g
Tubers, also called sunchokes, are high in both inulin and FOS.
Garlic
Both
9–16 g
Raw cloves provide the most; supports gut health via prebiotic effects.
Onions
Both
1.1–7.5 g (raw pulp)
Rich in FOS, red onions and shallots are particularly high.
Leeks
Inulin
3–10 g
Bulbs and leaves have a milder flavor than onions.
Asparagus
Inulin
2–3 g (raw)
Spears: Cooking may reduce levels slightly.
Dandelion Greens
Inulin
9.6 g (raw)
Leaves; bitter greens are often used in salads.
Bananas
Inulin
0.3–0.7 g (raw)
Slightly unripe (green) bananas are best.
Wheat
Both
1–3.8 g
Whole grains; bran is richest.
Burdock Root
Both
High (not quantified)
Root vegetable; used in teas and stir-fries.
Lentils
FOS
Moderate (not quantified)
Legumes also provide oligosaccharides.
Red Cabbage
FOS
Moderate (not quantified)
Fermented forms (e.g., sauerkraut) enhance benefits.
To maximize intake, aim for a variety of these foods daily (e.g., 5–10g total prebiotics).
Note that high doses may cause bloating in sensitive individuals, so start low.
You may wonder what the dietary sources of short-chain fatty acids (SCFAs) are, since they are so important in promoting overall health and longevity.
Short-chain fatty acids (SCFAs)—primarily acetate, propionate, and butyrate—are mostly produced endogenously by gut bacteria through the process of dietary fiber fermentation.
However, small amounts are available directly from certain foods.
Direct dietary sources provide limited quantities, often absorbed in the upper gut rather than reaching the colon for full benefits, so combining them with fiber-rich foods is ideal for optimal SCFA levels.
Below, sources are categorized as direct (naturally containing SCFAs) or indirect (fiber/prebiotic foods that promote SCFA production via fermentation).
Direct Sources (Foods Naturally Containing SCFAs) These include dairy products (from milk fats) and fermented items (where bacteria produce SCFAs during processing).
Amounts are modest (e.g., butter has ~3-4% butyrate by fat weight).
Dairy Products:
Butter and ghee: High in butyrate.
Cheese (e.g., hard varieties like Parmesan, pecorino): Contains butyrate and propionate.
Full-fat yogurt and milk (cow, goat, sheep): Provide butyrate.
Fermented Foods (SCFAs produced during fermentation):
Sauerkraut, kimchi, and some pickles: General SCFAs, including butyrate.
Kefir: SCFAs via fermentation.
Tempeh: Butyrate and other SCFAs.
Other:
Vinegars: Primarily acetate.
Some alcoholic beverages (e.g., certain wines or beers): Acetate.
Indirect Sources(Fiber-Rich Foods for Gut Production of SCFAs) These non-digestible carbs (e.g., resistant starch, inulin, pectins) are fermented by gut microbes to generate SCFAs, making up the bulk of intake (~90-95% of colonic SCFAs).
Aim for 25-30g fiber daily from a variety of plant sources.
Whole Grains and Cereals: Oats, barley, brown rice, whole wheat, rye. Brown rice and whole wheat pasta (cooked and cooled for resistant starch)
Legumes and Pulses: Beans (e.g., chickpeas, black beans), lentils, peas.
Vegetables: Asparagus, broccoli, carrots, onions, garlic, leafy greens, potatoes (cooked and cooled for resistant starch).
Nuts and Seeds: Flaxseeds, chia seeds.
Other:
– Resistant starches like cooled rice or cornmeal;
– Polyphenol-rich items (e.g., green tea, cocoa, dark chocolate, dark-skinned fruits, and dark leafy greens) that support SCFA-producing bacteria.
For maximum benefits, focus on indirect sources through a varied, plant-heavy diet, as they yield the most SCFAs in the colon. EAT THE RAINBOW!
Supplements exist but are less effective than food-based approaches.
Sample Daily Meal Plan for Promoting SCFAs
To support gut health and SCFA production, aim for 30–40g of dietary fiber daily from diverse plant sources like whole grains, legumes, fruits, and vegetables.
This sample plan provides approximately 37g of fiber and incorporates SCFA-promoting foods (e.g., brown rice for resistant starch, fruits for pectins, and vegetables for oligosaccharides).
It’s balanced for ~2,000 calories; adjust portions as needed. Focus on gradual increases to avoid digestive discomfort.
Breakfast (9g fiber)
Muesli (whole grain oats with nuts and seeds) served in milk with a drizzle of honey.
SCFA boost: Oats’ beta-glucan ferments into butyrate.
Beef curry (lean beef with onions, tomatoes, and turmeric, curry spices) served with brown rice.
Side salad of mixed greens with onions and tomatoes, avocado, and a lemon vinaigrette (olive oil 6 tbsp, mustard 1 tsp, lemon juice 4 tsp, lemon zest 1 tsp, honey 1 tsp, salt, pepper ).
SCFA boost: Brown rice’s resistant starch yields acetate and butyrate.
Afternoon Snack (2g fiber)
Plain low-fat yogurt with nuts
SCFA boost: Fermented dairy provides minor direct SCFAs and feeds beneficial bacteria.
Dinner (10g fiber)
Chicken risotto made with barley, mixed vegetables (carrots, peas, zucchini), and herbs.
SCFA boost: Vegetables and grains promote diverse fermentation for all major SCFAs.
Evening Snack (2g fiber)
A handful of berries (e.g., strawberries or blueberries).
Tips: Drink plenty of water (8+ cups/day) to aid digestion.
This plan draws from evidence showing high-fiber diets elevate plasma SCFAs like acetate and propionate within days.
For variety, swap in other sources like lentils or kiwi from the list above.
Consult a doctor for personalized advice, especially with gut conditions.
Read more about the critical role of SHORT-CHAIN FATTY ACIDS
What are Short-Chain Fatty Acids (SCFAs) and what role do they play in our overall health?
They play an enormous role in our health, encompassing gut health, metabolism, immunity, cardiovascular health, musculoskeletal health, neurological health, and mental health and wellbeing. Short-chain fatty acids (SCFAs), mainly acetate, propionate, and butyrate, are produced by our gut microbiota in the process of fermenting dietary fibers. Certain foods. provide small amounts of these amazing SCFAs
Direct dietary sources provide limited quantities, often absorbed in the upper gut rather than reaching the colon for full benefits, so combining them with fiber-rich foods is ideal for optimal SCFA levels.
Short-chain fatty acids influence health through G-protein-coupled receptors (e.g., GPR41/43), histone deacetylase (HDAC) inhibition, and systemic signaling.
Key Health Benefits of SCFAs Recent reviews highlight SCFAs’ roles in multiple systems, with emerging evidence from 2024–2025 studies emphasizing therapeutic applications.Gut Health
SCFAs fortify the intestinal barrier, suppress inflammation, and combat disorders like inflammatory bowel disease (IBD) by modulating Toll-like receptors (TLRs) and NLRP3 inflammasomes.
Butyrate promotes epithelial repair and mucus production, while acetate and propionate enhance antimicrobial defenses. They also reduce colon cancer risk via apoptosis induction in malignant cells.Metabolic Health
SCFAs improve insulin sensitivity, curb obesity, and alleviate metabolic syndrome by activating AMPK, boosting GLP-1/PYY for appetite control, and enhancing mitochondrial function. Propionate and butyrate specifically mitigate hepatic steatosis and dyslipidemia.
Clinical trials show SCFAs increase energy expenditure and reduce food intake.Immune and Antiviral Health
SCFAs drive anti-inflammatory responses, promoting regulatory T cells (Tregs) and IL-10 while curbing pro-inflammatory cytokines.
They act as antiviral mediators by enhancing interferon responses and barrier integrity against pathogens.
In IBD, they inhibit innate immune overactivation.Cardiovascular Health
SCFAs lower cholesterol absorption, reduce atherosclerosis via Treg expansion, and regulate blood pressure through Olfr78 receptor activation. Propionate decreases LDL levels and vascular inflammation.Neurological and Mental Health Via the gut-brain axis
SCFAs dampen neuroinflammation, reduce microglial activation, and lower apoptosis in models of Alzheimer’s and depression.
Supplementation decreases cortical inflammatory markers and improves cognitive outcomes.
SCFAs are a promising therapy for Parkinson’s, Alzheimer’s Diseases, Dementia, and Multiple Sclerosis.Skin and Aging Health
SCFAs support the gut-skin axis, modulating inflammation to promote barrier function, collagen synthesis, and anti-aging effects.
They link microbiome health to reduced skin disorders and delayed senescence.Cancer Prevention
SCFAs reverse cancer-linked epigenetic changes, inhibit tumor progression, and boost immunotherapy efficacy by inducing apoptosis and autophagy.
High-risk individuals may benefit from targeted SCFA administration to prevent epigenetic shifts.Other Benefits
SCFAs exhibit broad antimicrobial effects, protect against bone loss, and enhance muscle maintenance.
While generally beneficial, optimal intake via fiber-rich diets is recommended to avoid imbalances.
Read about Dietary sources of Short-Chain Fatty Acids – SCFAsRead more about the important role of SHORT-CHAIN FATTY ACIDS
Short-chain fatty acids (SCFAs), such as acetate, propionate, and butyrate, exhibit broad therapeutic potential across multiple disease categories, including neurodegenerative and demyelinating conditions.
They primarily act through mechanisms like G-protein-coupled receptor (GPCR) activation (e.g., GPR41/43/109A), histone deacetylase (HDAC) inhibition, anti-inflammatory effects (e.g., Treg promotion, NF-κB suppression), and metabolic regulation (e.g., AMPK activation for lipid/glucose homeostasis).
Comprehensive Therapeutic Applications of SCFAs Across Diseases
Short-chain fatty acids (SCFAs) demonstrate versatile therapeutic potential in a wide array of conditions, including neurodegenerative, autoimmune, metabolic, and gastrointestinal disorders.
Below is a summarized table of key diseases or conditions where SCFAs alleviate symptoms or show disease-modifying effects, based on recent reviews and studies.
These are grouped by category for clarity, with brief mechanisms, evidence levels noted, and links to clinical studies.
Category
Disease/Condition
Therapeutic Role/Mechanism
Key Evidence
Example Clinical Trial Link
Neurodegenerative
Alzheimer’s Disease
HDAC inhibition promotes microglial M2 shift, enhances Aβ phagocytosis/autophagy, upregulates BDNF for synaptic repair; restores BBB integrity via ZO-1/claudins.
APP/PS1 mouse models show plaque reduction (20–30%) and cognitive gains (MMSE +15–25%); 2025 RCTs in MCI confirm inflammation ↓ via FFAR2/3.
Suppresses α-syn aggregation via C/EBPβ/autophagy, modulates microglia (GPR109A/NF-κB inhibition), boosts GLP-1 for neuroprotection; restores gut barrier to curb L-dopa resistance.
MPTP models and 2025 pilots (tributyrin) report UPDRS ↓15–30% and motor improvements; FMT restores SCFAs, alleviating inflammation.
Epigenetic regulation (HDAC inhibition) modulates Aβ/tau pathologies; anti-inflammatory effects via Treg promotion and NLRP3 suppression; enhances brain metabolism and BDNF for cognitive function.
Preclinical AD models (most common dementia subtype) show synaptic repair and cognition ↑; 2025 reviews highlight gut-brain axis modulation as translational target.
SCFAs primarily alleviate symptoms and slow progression rather than cure; integration with diet/prebiotics/FMT enhances efficacy.
Consult a professional for the application.
SCFAs do not “cure” these conditions but show promise in alleviating symptoms, slowing progression, or enhancing standard therapies (e.g., via supplementation, prebiotics, or FMT (fecal transplant)).
Efficacy varies by SCFA type (butyrate is the most versatile), dose (500–2000 mg/day), and delivery (e.g., colon-targeted prodrugs).
Ongoing 2025 trials emphasize precision approaches, with the strongest evidence in metabolic and GI disorders.
Consult healthcare providers for personalized use.
Short-chain fatty acids (SCFAs), including acetate, propionate, and butyrate, emerge as promising therapeutic agents in Alzheimer’s disease (AD).
Short-chain fatty acids are produced by gut microbiota fermentation of dietary fibers.
These SCFAs play a complex role in Alzheimer’s disease (AD) through the microbiota-gut-brain axis.
AD patients exhibit gut dysbiosis with reduced SCFA-producing bacteria (e.g., Faecalibacterium prausnitzii, Roseburia spp.), leading to altered circulating SCFA levels—typically elevated acetate and valerate but decreased butyrate—which correlate with amyloid-β (Aβ) deposition, tau pathology, neuroinflammation, and cognitive decline.
SCFAs modulate AD progression by influencing microglial activation, blood-brain barrier(BBB) integrity, and synaptic plasticity, though effects can be beneficial (e.g., anti-inflammatory) or detrimental (e.g., impaired Aβ phagocytosis) depending on concentration, disease stage, and context.Recent 2024–2025 studies emphasize the SCFAs-microglia pathway as a therapeutic target, with preclinical evidence supporting microbiome modulation to restore SCFA homeostasis and slow neurodegeneration.
Key Mechanisms SCFAs exert dual effects in AD via epigenetic, signaling, and metabolic pathways, primarily targeting microglia—the brain’s resident immune cells that drive neuroinflammation and Aβ/tau pathology.
Epigenetic Regulation:
Butyrate and propionate inhibit histone deacetylases (HDACs), promoting hyperacetylation (e.g., H3K9, H3K18) that suppresses NF-κB translocation and pro-inflammatory genes (IL-1β, TNF-α, COX-2), shifting microglia from M1 (pro-inflammatory) to M2 (anti-inflammatory) phenotypes.
In APP/PS1 mice, oral acetate administration for 4 weeks upregulated GPR41 in Aβ-stimulated BV-2 microglia, inhibiting HDAC-related pathways and reducing inflammatory markers.
Sodium butyrate induced hyperacetylation at H3K9 and H3K18 sites in LPS-stimulated BV-2 microglia. In AD mouse models, sodium butyrate ameliorates synaptic plasticity impairment by inhibiting neuroinflammation via HDAC inhibition.
Receptor-Mediated Signaling: SCFAs bind G-protein-coupled receptors (FFAR2/3, GPR109A) on microglia, inhibiting TLR4/NF-κB and ERK/JNK pathways, reducing ROS/NO production, and enhancing phagocytosis or autophagy for Aβ clearance. Over 60% of hippocampal FFAR3 expression co-localizes with activated microglia. In APP/PS1 mice, acetate upregulated GPR41 in BV-2 microglia, inhibiting phosphorylation of NF-κB p65, ERK, and JNK, and reducing COX-2 and IL-1β levels. Butyrate reduced Aβ-induced CD11b and COX-2 in BV-2 microglia and inhibited NF-κB p65 phosphorylation. Knockout of GPR41/43 accelerated cognitive decline and impaired hippocampal neurogenesis in 5×FAD mice, but SCFAs intake reversed this by upregulating defensive genes (e.g., B2m, Fgl2, H2-K1) and antigen presentation pathways.
Metabolic Reprogramming: SCFAs restore tricarboxylic acid (TCA) cycle flux and mitochondrial function in microglia, balancing energy and curbing inflammasome (NLRP3) activation, which exacerbates synaptic loss in AD.
Gut-derived 13C-acetate can reach the brain and be metabolized by microglia into TCA cycle intermediates (e.g., citrate, α-ketoglutarate, fumarate, malate, succinate), thereby restoring the mitochondrial dysfunction observed in germ-free mice. In 5×FAD mice, acetate inhibited phagocytosis by inducing cytokine expression, exacerbating Aβ burden, and increased mitochondrial activity, ROS production, oxidative phosphorylation, and membrane potential in Aβ-phagocytosing microglia. Acetate improved TCA cycle flux by stimulating short-chain CoA metabolism and increasing acetyl-CoA levels, reducing microglial reactivity. Butyrate reversed FXN depletion-induced mitochondrial oxidative capacity loss via GPR109A, stimulating the itaconate-Nrf2-GSH pathway and reducing ROS.
Indirect Effects via Gut-Brain Axis: Circulating SCFAs influence peripheral immunity (e.g., Treg/Th17 balance) and vagal signaling, reducing gut permeability and systemic translocation of inflammatory signals to the brain. Propionate pre-treatment reduced peripheral Th17 infiltration and IL-17A levels, decreasing microglial activation in perioperative cognitive dysfunction models relevant to AD. FFAR2 knockout in myeloid cells downregulated microglial inflammatory genes.
SCFAs promoted Treg generation in the spleen, affecting microglial cytokine release. In 5×FAD mice, peripheral immune pathways mediated SCFAs’ effects on microglial transcriptome and neurogenesis. Elevated acetate may worsen Aβ burden by impairing microglial metabolism, while butyrate supports barrier integrity and BDNF expression.
SCFAs suppress pro-inflammatory cytokines (IL-1β, MCP-1, TNF-α) and reduce THP-1 phagocytosis; acetate reverses LPS-induced phospholipase C β1/COX-1/COX-2 and reduces TNF-α/IL-6 in astrocytes via p38 MAPK/NF-κB downregulation, increasing IL-4 via TGF-β1/H3K9 acetylation;
Butyrate inhibits COX-2 in Aβ-microglia via NF-κB.
Evidence from Preclinical and Clinical Studies
Studies reveal context-dependent SCFA effects, with 2025 cross-sectional data confirming AD-specific plasma signatures.
Below is a summary of key 2024–2025 findings:
Study Type/Source
Key Findings
Model/Population
Outcomes/Implications
Cross-Sectional Observational (PMC, Jun 2025)
Elevated plasma acetate/valerate and reduced butyrate in CI-AD (n=28) vs. controls (n=10) and non-AD impairment (n=29); valerate ratios positively correlate with amyloid PET (rho=0.35–0.59) and GFAP/NFL (rho=0.45–0.59). Acetate distinguishes CI-AD from non-AD (AUC=0.954).
Human cohorts (n=67)
SCFAs as biomarkers for AD differential diagnosis; excess acetate links to inflammation, butyrate depletion to pathology.
Review: SCFAs-Microglia Pathway (J Neuroinflammation, May 2025)
Butyrate suppresses Aβ-induced microglial activation (CD11b/COX-2 ↓) via HDAC/NF-κB inhibition; acetate reduces LPS-ERK/JNK in BV-2 cells. GPR41/43 KO worsens hippocampal neurogenesis; SCFAs reverse via defensive genes (B2m, Fgl2 ↑). Dual effects: germ-free models show SCFAs ↑ APOE, impair Aβ phagocytosis.
APP/PS1, 5xFAD mice; BV-2/in vitro microglia
Highlights dose/stage dependency; supports targeted modulation to enhance M2 shift and clearance.
Propionate ↓ microglial phagocytosis of fibrillar Aβ, maintains homeostatic phenotype without M2 shift.
Aβ-induced IMG microglia (in vitro)
Cautions against indiscriminate supplementation; low doses may impair clearance in early AD.
Microbiota-FMT (Mol Nutr Food Res, cited 2025)
Clostridium butyricum colonization ↑ butyrate, inhibits microglial activation via GPR43 in APP/PS1.
APP/PS1 mice
FMT boosts SCFA-producers for anti-inflammatory effects.
Human evidence is emerging:
Salivary acetate/propionate ↑ in AD, correlating with periodontal risk; plasma SCFAs associate with brain acetate uptake in MCI.
Therapeutic Applications SCFAs offer adjunctive strategies to target early AD dysbiosis, with 2025 reviews advocating precision interventions to leverage beneficial effects while mitigating risks like impaired phagocytosis.
Supplementation: Sodium butyrate (500–2000 mg/day) or prodrugs (e.g., tributyrin) restore levels, inhibit HDACs, and improve cognition in models;
Clinical pilots explore oral dosing for MCI (Mild Cognitive Impairment)
Prebiotics/Probiotics: Inulin or galacto-oligosaccharides (5–10 g/day) enrich SCFA-producers, reducing microglial senescence and plaques (e.g., 20–30% inflammation ↓ in aged models).
Strains like Bifidobacterium breve or Roseburia hominis via psychobiotics enhance butyrate, supporting synaptic repair.
FMT and Diet: Fecal transplants from healthy donors ↑ SCFAs, alleviate neuroinflammation in AD models; high-fiber Mediterranean diets elevate circulating levels, correlating with slower progression.
Novel Targets: Microglia-specific FFAR2/3 agonists or colon-targeted delivery (e.g., acylated starch) optimize brain penetration; combined with anti-Aβ therapies for amyloid-positive patients.
Doses are safe (up to 4 g/day), but variability from microbiome baseline requires multi-omics personalization. Challenges include dual effects and BBB (blood-brain barrier) crossing;
Ongoing 2025 trials (e.g., prebiotic RCTs in MCI) aim to validate 15–25% cognitive gains.
SCFAs hold transformative potential for AD prevention, bridging gut modulation to neuroprotection.
Gut dysbiosis, marked by diminished microbial diversity and imbalanced bacterial composition, is a hallmark of Alzheimer’s disease (AD)
AD often emerges in prodromal stages like mild cognitive impairment (MCI) and contributes to pathogenesis through the microbiota-gut-brain axis.
AD patients show consistent reductions in short-chain fatty acid (SCFA)-producing taxa and enrichments in pro-inflammatory genera, correlating with amyloid-β (Aβ) plaques, tau hyperphosphorylation, neuroinflammation, and cognitive metrics (e.g., MMSE scores).
2024–2025 meta-analyses and cohorts reveal geographic and stage-specific variations, with dysbiosis driving “leaky gut,” metabolite dysregulation, and immune activation that exacerbate BBB (blood-brain barrier) permeability and microglial priming.
This supports a gut-first hypothesis, where dysbiosis precedes and amplifies AD progression, offering targets for early microbiome-based interventions.
Microbial Alterations in AD Meta-analyses indicate inconsistent α-diversity reductions (significant in AD but not always MCI), with β-diversity shifts reflecting compositional changes.
Key patterns involve depleted anti-inflammatory/SCFA-producers and elevated opportunistic pathogens, with fecal SCFA levels (e.g., butyrate) often decreased by 20–40%.
Pattern
Key Taxa Changes
Correlations & Evidence
Reduced Diversity & Beneficial Depletion
↓ α-diversity (Shannon/Simpson indices in AD); ↓ Firmicutes, Blautia, Roseburia, Faecalibacterium prausnitzii, Lachnospiraceae, Rikenellaceae, Clostridiaceae; ↓ Coprococcus comes, Odoribacter splanchnicus, Roseburia intestinalis (monotonic decline with CDR/GDS-FAST severity)
Lower SCFAs; associates with amyloid PET (rho=0.35–0.59), GFAP/NFL (rho=0.45–0.59), cognitive decline (MMSE ↓), and NPS/depression; observed in meta-analyses (n=805) and Spanish cohort (n=97).
Increased permeability/inflammation (fecal calprotectin ↑, LPS ↑); links to Aβ/tau pathology, microglial activation; U.S.-specific ↑ Bacteroides/Alistipes vs. ↓ in China; gradient in AD > MCI.
Evidence from Preclinical and Clinical Studies 2024–2025 research highlights causality via FMT models and multi-omics, with human cohorts (n>1,000) confirming biomarkers.
Study Type/Source
Key Findings
Model/Population
Outcomes/Implications
Meta-Analysis (Alzheimers Dementia, Dec 2024)
Complex dysbiosis-cognition link; reduced beneficial taxa correlate with impaired function.
11 studies (n=805)
Dysbiosis as modifiable risk; influences amyloid/inflammation.
Cohort Characterization (PMC, Sep 2025)
No sig diversity diff, but SCFA-producer declines with severity; Parabacteroides distasonis ↑ with depression/NPS.
Spanish elderly (n=97: HC/MCI/AD)
Mediterranean lifestyle buffers; taxa as cognitive biomarkers.
Gut dysbiosis is characterized by reduced microbial diversity and shifts in bacterial composition.
It is a prominent feature in Parkinson’s disease (PD), often preceding motor symptoms by years and contributing to disease initiation and progression via the microbiota-gut-brain axis.
PD patients exhibit consistent alterations, including depletion of short-chain fatty acid (SCFA)-producing bacteria and enrichment of pro-inflammatory taxa, which correlate with gastrointestinal symptoms (e.g., constipation), non-motor issues (e.g., depression, sleep disturbances), and motor severity (e.g., UPDRS scores).
These changes are influenced by factors like disease duration, medications, diet, and geography, with emerging evidence from 2024–2025 studies supporting a “gut-first” model.
In this model, dysbiosis drives α-synuclein pathology, neuroinflammation, and dopaminergic neuron loss.
Longitudinal profiling and fecal microbiota transplantation (FMT) models underscore causality, positioning dysbiosis as a modifiable target for early intervention.
Microbial Alterations in PD Meta-analyses and cohort studies reveal reproducible patterns, though alpha diversity reductions are often non-significant due to confounders.
Key shifts include decreased beneficial, anti-inflammatory genera and increased opportunistic pathogens, with fecal short-chain fatty acids (SCFA) levels (e.g., butyrate) reduced by 20–50%.
Lower SCFA production correlates with constipation, disease progression (e.g., Hoehn & Yahr stage), and motor/non-motor symptoms (NMSS scores ↑); observed in fecal/ileal samples from PD (n=44) vs. HC (n=21).
Increased gut permeability and inflammation (fecal calprotectin ↑); links to α-syn aggregation and motor deficits (e.g., beam walking time ↑ in FMT models); ileal SFB erosion in PD mice/patients.
Other Shifts
↓ Segmented filamentous bacteria (SFB) in ileum; variable Bifidobacterium (depleted in ileal biopsies)
Key Mechanisms: Dysbiosis initiates a cascade from gut to brain, with bidirectional gut-brain signaling via vagus nerve, metabolites, and immune cells.
Increased Intestinal Permeability (“Leaky Gut”):
Depleted Prevotella impairs mucin production, thinning colonic mucus, and downregulating tight junctions (e.g., ZO-1, occludin).
Sulfate-reducing bacteria (e.g., Bilophila) produce excess H₂S, degrading mucus.
Reduced SCFAs weaken barriers, allowing pathobionts/LPS translocation; TNF-α internalizes ZO-1, elevating fecal calprotectin.
In PD ileum, this correlates with CD11b+ immune cell influx and pro-inflammatory cytokines (TNF, IL-6, IL-8).
Neuroinflammation and Immune Dysregulation:
Pro-inflammatory taxa (e.g., Enterobacteriaceae) activate TLR4/NF-κB, elevating cytokines (IL-17, IL-1β) and shifting Th17 from homeostatic to inflammatory phenotypes (↓ CD4+/IL-17+ cells, ↑ CD8+ IL-17).
SFB erosion reduces Th17 induction, promoting chronic gut inflammation that propagates systemically (↑ plasma IFNγ, IL-6) and to brain (microglial Iba1+/Trem2+ activation, NLRP3 inflammasome).
This exacerbates dopaminergic loss in the substantia nigra (SN; ~30% TH+ neurons ↓).
α-Synuclein Aggregation and Propagation:
Pathobionts like E. coli (curli proteins) and Proteus mirabilis (hemolysin A) induce ENS α-syn misfolding/phosphorylation (p-α-syn ↑), propagating caudo-rostrally via vagus to dorsal motor nucleus (DMV) and SN. Dubosiella disrupts lysosomal function via branched-chain amino acid buildup.
TMAO from dysbiosis promotes aggregation/NF-κB. p-α-syn correlates with mitochondrial fragmentation (TOM20+ ↓) and precedes motor deficits in FMT models (week 3 onset).
Oxidative Stress and Mitochondrial Dysfunction:
Dysbiosis reduces antioxidants (e.g., via ↓ NMNAT2/NAD+), upregulates NOX4/ROS, and inhibits Nrf2.
Bacterial PAMPs/mitochondrial DAMPs (e.g., cardiolipin) activate caspase-1/IL-1β;
Sleep deprivation worsens via adenosine-NOX4. Leads to SN ATP ↓ (~52% striatal dopamine reduction) and BBB (Blood-Brain Barrier) leakage (IgG+ leaks).
Neurotransmitter and Metabolite Imbalance: ↑ Tyrosine decarboxylase in gut bacteria converts L-dopa prematurely, reducing efficacy.
↓ SCFAs compromise BBB;
↑ Secondary bile acids/TMAO impair autophagy/mitochondria.
Disrupts dopamine/serotonin synthesis, linking to hyposmia and mood symptoms.
Evidence from Preclinical and Clinical Studies 2024–2025 research emphasizes ileal dysbiosis and FMT causality, with human cohorts (n>60) and mouse models replicating PD-like pathology.
Study Type/Source
Key Findings
Model/Population
Outcomes/Implications
Human Cohort (Fecal/Ileal) (Mol Neurodegener, Oct 2024)
↑ Lactobacillus/Streptococcus, ↓ Faecalibacterium/Roseburia; ileal SFB ↓, Enterobacteriaceae ↑; correlates with gut inflammation (ZO-1 ↓, cytokines ↑) and motor scores.
PD patients (n=44 fecal, n=2 ileal) vs. HC (n=21)
Supports gut-first model; ileal biomarkers for early detection.
FMT Mouse Model (Front Neurosci, Jun 2025)
PD-FMT induces dysbiosis (↓ Roseburia, ↑ Akkermansia), leaky gut, α-syn spread, SN neuron loss; reversed by HC-FMT.
Probiotics/Synbiotics: L. plantarum PS128/DP189, B. breve CCFM1067, C. butyricum reduce α-syn/ROS via GLP-1/miR-155; pilots: motor/QoL improvements (8–12 weeks).



{kind=link}
{kind=link}
{kind=link}